IV. Размножение растений
IV. Размножение растений
Размножение растений в культуре совершается весьма различными способами, которые, однако, несмотря на разнообразие в исполнении, относятся к двум главным категориям; а) половое размножение – семенами, являющееся в результате оплодотворения и ставящее новое растение в зависимость от родителей и их способности передавать свойственные им особенности; b) бесполое размножение, – клубнями, почками, ветками, усами, корнями, делением, отпрысками, прививкой и проч. Бесполое размножение всегда или за весьма редкими исключениями верно передает все особенности материнского растения; например, от прививки садового сорта Антоновского яблока всегда получается этот сорт, между тем как при половом размножении сорт может видоизмениться, – в противоположность настоящим видам, например, дикому яблоку, которое редко отклоняется от типичной формы.
А. Половое размножение
1. Оплодотворение растений
При образовании семян покрытосемянных (Angiospermae) растений участвуют два элемента: мужской и женский. Для образования нового организма – зародыша, соединяются две микроскопические клеточки, образуя новое «неделимое». Самый акт опыления и оплодотворения совершается таким образом: зрелая пыльца, выступающая при растрескивании пыльника, переносится или случайно, или особыми приспособлениями растений, или насекомыми, или умышленно культиватором – на рыльце, т. е. на женский орган цветка. Зрелость рыльца, т. е. готовность его к восприятию пыльцы обнаруживается тем, что на нем выделяется слизистая сахаристая жидкость, к которой прилипают крупинки цветочной пыли. Жидкость эта представляется некоторым образом почвою, на которой прорастает пыльца; при отсутствии этой жидкости прорастание пыльцы не происходит, а следовательно, и оплодотворение невозможно. Пыльца представляет микроскопические клетки, различной у различных растений формы, имеющие две оболочки – наружную и внутреннюю. На наружной, более толстой оболочке (exina) имеются утонченные места, через которые выступает внутренняя оболочка, (intina), в форме цилиндрической трубки при прорастании пыльцы. Прорастая, пыльцевая трубочка проникает через нежную ткань рыльца в рыхлую ткань или полость столбика, следуя дальше в полость завязи. Здесь пыльцевые трубочки направляются к семяпочкам, следуя у большинства высокоразвитых Angiosper-mae по направлению к ткани, ведущей прямо к пыльцевходу (micropyle), через который пыльцевая трубочка проходит в зародышевый мешок. Иногда пыльцевая трубка не направляется к micropyle, а проникает в зародышевый мешок иными путями, проходя через покровы семяпочки или через семяножку (халадзу). Еще пребывая на рыльце, ядро пыльцы, представляющей собою одну клетку, делится, образуя две клетки – вегетативную и генеративную, которые, однако, находясь в общей оболочке, не разъединены друг от друга перегородкой. Во время роста пыльцевой трубки заключающееся в ней «генеративное ядро» (генеративной клетки) делится на два генеративных ядра, называемых «живчиками» или «сперматозоидами», которые постепенно спускаются на конец пыльцевой трубки и которые, во входе пыльцевой трубки в зародышевый мешок через микропиле, проникают в последний для оплодотворения.
К этому времени зародышевый мешок является уже совершенно развитым и готовым для процесса оплодотворения; в нем к этому времени имеем: 1) у микропиле три клеточки, составляющие так называемый «яйцевой аппарат» (яйцеклетку и две вспомогательные клетки – «синериды»), 2) в противоположном конце сходятся еще три клеточки, называемые «антиподами», содействующими, как полагают, питанию зародышевого мешка, частей и зародыша (Веттштейн) и 3) вторичное ядро «зародышевого мешка» (являющееся результатом слияния двух «полярных ядер»), которые, однако, к моменту оплодотворения часто еще не бывают слитыми, – тогда вместо одного ядра мы имеем их два. Вторичное ядро зародышевого мешка (или два полярных) расположены обычно в центре зародышевого мешка (см. рис. 15). Конец пыльцевой трубки проникает в зародышевый мешок обыкновенно между синергидами, почему последние и называются «вспомогательными клетками». К этому времени генеративное ядро пыльцевой трубки разрушается, а из двух вегетативных ядер («живчиков» или «сперматозоидов») одно проникает к яйцеклетке и сливается с ней в одно ядро, а другое направляется к вторичному ядру зародышевого мешка (или к двум рядом находящимся и не слитым еще полярным ядрам) и сливается с ним (или с обоими вместе полярными ядрами).
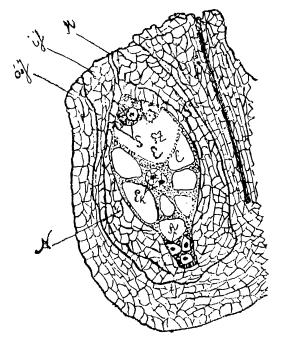
Рис. 15. Семяпочка подснежника (Scilla maritime): aJ – внешний покров семяпочки (интегумент); iJ – внутренний покров; M – микропиле; F – семяножка; N – nucellus; E – зародышевый мешок; S – вспомогательные клетки или синергиды; Ez – яйцеклетка;Ek – ядро зародышевого мешка, получающееся из слияния двух полярных ядер;A – антинозы (по Миэ)
Из оплодотворенной яйцеклетки развивается зародыш растения, а из оплодотворенного вторичного ядра зародышевого мешка (или двух полярных ядер) развивается белок – endospermum. В этом, собственно, и состоит акт оплодотворения у покрытосемянных растений, названный «двойным оплодотворением», часть открытия которого принадлежит нашему известному русскому ботанику проф. С. Г. Навашину.
У голосемянных (Gymnospermae) процесс оплодотворения происходит вполне аналогично указанному. Однако у них нет ни завязи, ни рыльца, ни столбика, а семяпочки с одним только покровом расположены на семенных чешуйках женского цветка на верхней стороне последних (поэтому растения этого рода и называются в отличие от покрытосемянных «голосемянными»). Покров семянопочки «интегумент» в верхней части его не вполне смыкается и, несколько выступая над ядром (nucellus), образует небольшое отверстие – микропиле (micropyle). Зародышевый мешок (см. рис. 16), заключенный глубоко в ткань семяпочки, называемой ядром (nucellus), представляет собою к моменту оплодотворения многоклеточное тело, в котором, на обращенном к micropyle конце, имеются два или много архегониев, заключающих по одной большой яйцеклетке. Готовые к оплодотворению яйцеклетки также отделены слоем ткани от образующейся под микропиле пыльцевой камеры.
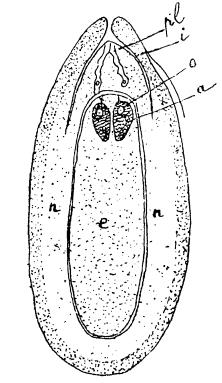
Рис. 16. Продольный разрез через семяпочку Pinus Laricio: pl – пыльцевая трубка; – семенная оболочка; n – nucellus, или ткань ядра семяпочки; е – эндосперм (или зародышевый мешок); а – архегоний; о – яйцеклетка (по Соulеr и Chamberlain)
Когда яйцеклетки готовы к оплодотворению, то из микропиле у хвойных выступает капелька жидкости, на которую обыкновенно и попадает летающая в воздухе пыльца хвойных во время цветения их. Как только пыльца попала на эту жидкость, последняя всасывается обратно; таким образом пыльца переносится в пыльцевую камеру, где она начинает прорастать в ткань ядра семяпочки и достигает яйцеклетки, образуя короткую пыльцевую трубку. При прорастании пыльцы в пыльцевой трубке обыкновенно образуются два генеративных ядра; у некоторых хвойных бывает и большее количество генеративных ядер, например, у Araucaria от 14 до 44. Эти ядра по пыльцевой трубке направляются к яйцеклетке, при чем одно из них сливается с яйцеклеткой – в результате чего развивается зародыш.
У мхов и папоротников, относимых нами, как и голосемянные, к архегониальным растениям, размножение производится бесполыми одноклеточными спорами. Из этих спор вырастают небольшие растеньица, производящие половые органы: женские – архегоний и мужские – anmepuОии. У папоротников, например, с которыми садоводам чаще всего приходится иметь дело, эти молодые растеньица, представляющие собою половое поколение, имеют большею частью вид небольших сердцевидных зеленых чешуек, называемых «предростками» – prothalium (гаметофит), которые прикреплены к почве тонкими бесцветными волосками, играющими роль корней. На нижней же стороне, обращенной к земле, у верхнего лопастного конца этого предростка развиваются архегонии, т. е. образования в виде бутылочки (колбочки), брюшная часть которой погружена в ткань предростка, а горлышко или шейка выступает наружу, образуя небольшой выступ. На дне этой колбочки находится одна только яйцеклетка. Ниже архегоний на той же пластинке образуются «сперматозоиды» в виде небольших округлых выростов отдельных леток предростка; в антеридиях образуются «сперматозоиды», имеющие большое количество жгутиков или ресничек, с помощью которых они свободно передвигаются в каплях воды, достигая таким образом архегоний и проникая во внутрь последних к яйцеклетке, с которой и сливается один из многих сперматозоидов, образуя зародыш. Зародыш этот, продолжая развиваться, образует новое молодое растение, на котором в дальнейшем, по мере его развития, развиваются споры одинаковой величины (равноспоровые папоротники); это так называемое бесполое поколение (спорофит), иногда развивающееся в огромные растения, является излюбленными растениями, культивируемыми у нас в оранжереях и теплицах (например, древовидные папоротники).
Кроме равноспоровых, мы имеем и разноспоровые папоротники, т. е. такие, которые образуют споры двоякого рода: макроспоры и микроспоры. Развивающиеся из этих спор молодые растеньица (предростки – гаметофиты) бывают раздельнополыми: из микроспор вырастают только мужские особи, образующие антеридии, а из макроспор – женские, несущие только архегонии. Как женские, так и особенно мужские предростки бывают значительно редуцированными по сравнению с предростками настоящих папоротников.
Самый процесс оплодотворения у последних проходит аналогично указанному выше у равноспоровых.
Половые органы у лиственных мхов развиваются на концах главных или коротких лиственных побегов мха, т. е. на взрослом растении (гаметофите). Антеридии и архегонии находятся или на одном и том же стебле вместе (однодомные), или же на разных стеблях (двудомные растения). После оплодотворения яйцеклетка начинает делиться и превращается в спорогоний (спорофит).
Спорофит во взрослом состоянии состоит из верхней части – коробочки – и нижней удлиненной – ножки, основание которой ущемлено в ткань стебля гаметофита. Таким образом гаметофит у мхов значительно сильнее развит, нежели спорофит. После созревания спор коробочки раскрываются, и споры, прорастая, дают начало развитию так называемой «протонеме», имеющей вид нитчатого, иногда значительно разветвленного или даже пластинчатого тела, из которого развиваются стебельки взрослого растения (гаметофита).
У водорослей половой процесс протекает различно: у низших водорослей половой акт состоит в слиянии двух равнозначных половых гамет, свободно движущихся в воде. Далее у некоторых водорослей оплодотворение состоит в слиянии или «копуляции» двух равноценных по форме и свойствам, свободно движущихся в воде гамет, но различных по величине: мужской – маленькой и женской – более крупной. Наконец, у высших водорослей мы имеем типичный половой процесс, свойственный по существу и высшим растениям, состоящий в слиянии подвижного мелкого живчика (сперматозоида), развивающегося в мужеском половом органе, называемом «спермагонием» (антеридием), с крупным пассивным яйцом («оосферой»), развивающимся в женском половом органе, называемом «оогонием». Половые органы водорослей являются одноклеточными, чем они резко отличаются от половых органов мхов и папортникообразных. Процесс оплодотворения у высших водорослей состоит в том, что живчики, двигаясь в воде и встречая женские половые органы, проникают в них через отверстия, имеющиеся в оогониях и достигая яйца, сливаются с ним. Хотя в оогоний может проникнуть много живчиков, но всегда сливается с женским только один. Таким образом происходит оплодотворение. Оплодотворенное яйцо водорослей именуется «ооспорой», а самый процесс оплодотворения «оогамией» в отличие от «изогамии», когда оплодотворенная клетка получилась путем слияния двух одинаковых половых клеток. Ооспора водорослей бывает одета плотной, толстой и непроницаемой оболочкой и потому может переносить различные неблагоприятные условия в покоящемся состоянии. Попадая в благоприятные условия, ооспора или непосредственно прорастает в водоросль, начинающую затем размножаться бесполыми зооспорами, или содержимое ооспоры повторно делится несколько раз, образуя бесполые зооспоры, которые, поплавав некоторое время, оседают, втягивают реснички, высачивают оболочку и прорастают, образуя водоросль.
Половой процесс у грибов выражен крайне слабо и в настоящем его виде наблюдается только у некоторых низших, развивающих «антеридии» и «оогонии», причем в антеридиях сперматозоиды не образуются, а оплодотворение яйцеклетки происходит таким образом, что антеридий дает небольшой трубчатый вырост, проникающий в полость оогония, где, соприкасаясь с яйцеклеткой, ядро антеридия переходит непосредственно в протоплазму яйцеклетки, сливаясь с последней – в результате чего образуется ооспора. Однако и у этих грибов далеко не всегда происходит оплодотворение; у высших грибов настоящий половой процесс совершенно отсутствует, и они размножаются или бесполым путем при посредстве спор и конидий, или же чаще всего вегетативным путем при посредстве мицелия (например, шампиньон – Psalliota campestris).
Между актом опыления, т. е. переносом цветочной пыльцы на рыльце, и актом оплодотворения, т. е. слиянием содержимого пыльцевой трубки с яйцеклетками, проходит более или менее продолжительное время, – от нескольких часов до полудня, вообще редко более суток, за исключением некоторых хвойных растений, напр., сосны, можжевельника и прочих растений с двухлетним плодосозреванием, у которых опыление совершается в начале июня, а оплодотворение спустя целый год. У некоторых растений столбики, чрез которые прорастают пыльцевые трубки, имеют (например, у Datura и Mirabilis) 3–4 дюйма длины; у мака рыльце сидит непосредственно на плоднике; у хвойных ни рыльца, ни столбика, ни завязи не существует. Но все такие различия – нормальные явления для данных растений и не изменяют физиологического значения оплодотворения и его действия.
2. Действие оплодотворения. Плодородие и бесплодие
Оплодотворение всего удачнее совершается между растениями одного и того же естественного вида и, как доказано опытами, удачнее между двумя растениями или двумя обоеполыми цветками одного и того же растения, нежели при самоопылении обоеполых цветков; есть некоторые луковичные растения, которые даже скорее принимают опылителя близкого к ним вида, чем их собственного. Также различные разновидности одного вида весьма легко оплодотворяются между собою. От посаженных вместе двух сортов кочанной капусты, коломенской и ульмской, я получил гораздо большее число помесей, чем чистопородных растений. За пределами вида затруднительность оплодотворения возрастает, и во многих случаях оно невозможно; от перекрестного опыления земляники и клубники мне при нескольких опытах не удалось получить семян, между тем как это вовсе не трудно с другими видами земляники, яблонями, смородиною и проч. Между растениями различных родов, – напр., между рябиною и грушею, малиною и земляникою – оплодотворение весьма редкое явление; известны лишь немногие такие гибриды между злаками, рододендронами и геснериевыми.
При соединении, для создания зародыша, двух половых клеточек одного вида обычно всегда получается потомство таких же свойств, напр., от дуба такой же точно дуб. Растения, представляющие собою разновидности, например яблони, груши, крыжовник, смородина, наоборот – редко дают от семян растения тех же разновидностей: их потомство более или менее приближается к дикому виду, хотя нет сомнения, что такие растения через многочисленные генерации, при тщательном выборе производителей, могут сделаться столь же константными, как и различные разновидности огородных плодовых растений, каковы, например, дыни, арбузы, огурцы, помидоры и проч.
Далее – потомство от скрещивания двух видов обыкновенно представляет собою что-то среднее между ними (гибрид). Большинство разводимых у нас высокорослых древесных ив, вероятно, такие гибриды от белой и ломкой ивы. При помощи гибридизации или скрещивания видов достигнуто создание и улучшение многочисленных сортов огородных и садовых растений. Естественные виды обыкновенно устойчивы в культуре в течение довольно продолжительного времени; но, раз уже гибридизированные или впавшие в неустойчивость путем видоизменения, они дают многочисленные сорта, которых у земляники, яблони и роз в настоящее время существует по тысяче и более. При гибридизации слишком отдаленных между собою видов случается, что потомство теряет способность полового размножения, такие растения представляют собою бесплодные гибриды.
Гибридизация, однако, не единственная причина бесплодия, которое также может произойти от изменения культуры или условий существования вида. Яблоко гравенштейнер, груша бессемянка, жемчужный, египетский и рокамбольский лук, хрен и некоторые сорта картофеля и хмеля бесплодны или почти бесплодны, т. е. являются бессемянными растениями. Клубника, особенно на старых грядах, часто становится бесплодной (пустоцветом). Во многих случаях половая способность может быть восстановлена одною удачно выведенною половым путем генерациею; в других случаях бесплодные гибриды и разновидности размножаются бесполым путем: прививками, черенками, бульбами, делением и так далее. У клубники бесплодные растения бросают и разводят новые гряды с плодоносными растениями; даже усы от бесплодных растений становятся при хорошем уходе плодоносными, хотя ими для размножения следовало бы пользоваться только в крайнем случае.
3. Содействие естественному опылению
Удивительно, с какой верностью выполняется опыление в природе, даже в таких случаях, где оно на первый взгляд кажется невероятным. Я много лет сряду наблюдал женский экземпляр пятитычиночной ивы, который находился на расстоянии более версты от мужского экземпляра того же вида, но тем не менее ежегодно он приносил во множестве спелые семена. Собранные и высеянные осенью семена эти дали отличные всходы настоящего вида, хотя весьма возможно, что пчелы, которые произвели здесь опыление, переносили пыльцу и с других видов ив, но в этом случае постороннее опыление оказалось бессильным, как почти всегда, когда пыльца других видов переносится в смеси с пыльцою настоящего вида. Что опыление действительно производилось пчелами, доказывается тем, что покрытые кисеей ветви не дали семян.
В культуре, напр., в оранжереях, теплицах и парниках, растения находятся в искусственных условиях, и опыление здесь не так легко совершается. При раннем разведении, напр., огурцов, дынь и арбузов необходимо искусственно помогать опылению, перенося пыльцу на женские цветки кисточкою или перенося целые тычинки со спелою пыльцою, которую высыпают на рыльце; если возможно, то пыльники прямо кладут на рыльце и оставляют их лежать на нем. Некоторые плодоводы задают себе громадный труд опылять все отдельные цветки персиковых деревьев. Вообще деревья, привязанные к шпалерам и вследствие того лишенные всякого движения, часто страдают недостаточным опылением. Но есть и более простой способ способствовать ему: когда пыльники растрескиваются, тогда ударяют чем-либо о шпалеры; вследствие сотрясения цветочная пыль разлетается по всем направлениям и падает на рыльца. Эта манипуляция производится раза два-три в день во время цветения. Сухой и теплый воздух способствует созреванию и распылению цветня по воздуху; поэтому во время цветения не допускается ни опрыскивания, ни спертого воздуха. Умеренное движение воздуха весьма способствует опылению; ввиду этого в парниках и теплицах во время цветения дают по возможности более воздуха, конечно, не подвергая при этом цветы простуде или замерзанию. Слишком высокой температуры во время цветения также должно избегать; иначе случается, что пыльники, равно как и пестики, не развиваются нормально, а завядают раньше, чем совершится опыление. То же самое может случиться от слишком сильного нагрева солнца, почему полезно вообще защищать цветы от солнца легким отенением в продолжение нескольких часов пополудни.
Еще вреднее действует на опыление и оплодотворение темнота, сырость и холод. Цветы всегда первые изо всех органов растений страдают от низкой температуры, равно как и от слишком высокой.
Если имеют дело с двудомными растениями, каковы спаржа, облепиха, шпинат, конопля и им подобные, и желательно получить от них семена, то необходимо рядом с женскими экземплярами иметь и мужские, чтобы между ними совершалось опыление. В таких случаях свойства и качество семян столько же зависят от мужских, сколько и от женских растений и, следовательно, необходимо обращать такое же внимание на выбор первых, как и последних.
4. Производство искусственного опыления с целью гибридизации или скрещивания
Гибридизация – одно из могущественных средств к созданию новых и улучшению существующих сортов культурных растений. Чтобы вполне убедиться в справедливости этого, приведем пример: от садовой яблони нет видоизменения, которое было бы вполне выносливо к морозам в северных и даже средних губерниях; все сорта довольно часто более или менее страдают от морозов и иногда совершенно вымерзают. Есть, однако, два диких вида, сибирское и китайское яблоко (Pyrus baccata et prunifolia), которые вполне выносливы к северной зиме, но как плодовые деревья малоценны в их естественном состоянии, хотя и существуют видоизменения, употребляемые в пищу. Путем гибридизации этих лучших видоизменений с хорошими и более выносливыми садовыми сортами возможно создать целую группу улучшенных плодовых деревьев, способных переносить суровые климатические условия северных губерний. Смотря по желанию, при выборе производителей можно действовать и в других направлениях: например, в направлении улучшения качества, величины, плодородности, формы и окраски данного материала. Почти все сорта нашей крупноплодной земляники и разных ягодных кустарников созданы таким путем, но они созданы в западной Европе и потому не обладают тою выносливостью, какая была бы желательна для разведения их на севере. Для наших суровых условий необходимо было создать новые формы этих растений – более выносливые, скороспелые, доброкачественные и проч.
При производстве гибридизации между видами и при скрещивании сортов поступают различным образом, смотря по тому, с какими растениями имеют дело. Возьмем сперва:
а) однолетнее или двулетнее огородное растение – капусту, репу, редьку. Многие из них обладают хорошими качествами, но оставляют еще многого желать относительно общего сложения растения, напр., коломенская капуста; другой пример: ульмская капуста – превосходного сложения, но не так высока по качествам, не так бела, нежна и сладка, как коломенская. От скрещивания обеих получается (по опыту) весьма улучшенная ульмская капуста, но едва ли улучшенная коломенская. Для скрещивания требуется только посадить растения в смеси на одной гряде, и скрещивание совершается само собою во время цветения. Разумеется, для этой доли необходимо, чтобы оба сорта цвели одновременно, для чего обрезают как преждевременные, так и запоздавшие цветы. Несмотря, однако, на то что растения посажены в смеси, необходимо собирать семена каждого сорта отдельно.
b) В случае, если имеют дело с небольшими травянистыми растениями, которые, как, например, земляника, разводятся бесполым способом и от которых, следовательно, нет надобности получать большого количества семян, а лишь несколько семян известного происхождения (для получения новых сортов), то удобнее сажать выбранные растения в горшки рано весною и помещать таковые в оранжерею, в парник или даже на окошко в комнате, обращенной на восток. Цветы земляники, как и у многих других культурных растений, обоеполые – мужские и женские половые органы находятся во всех цветках; это обстоятельство и усложняет операцию. Необходимо устранить естественное опыление, для чего тычинки вырезаются раньше, чем раскроются пыльники, при помощи тонкого и острого пинцета. Тычинки у земляники многочисленны и очень мелки; они развиваются постепенно, и требуется большая осмотрительность, чтобы не оставить ни одной на дне цветка. На каждом цветочном стебельке оставляется только по одному или по два цветка – остальные удаляются еще до распускания.
Когда рыльца, которые у земляники тоже многочисленны, достигнут зрелости – покроются блестящею жидкостью (что можно наблюдать в лупу), то переносят на них пыльцу, которую можно собирать с тычинок за день перед опылением, или же, что даже удобнее, кладут на оплодотворяемый цветок другой цветок в обратном положении, со спелыми тычинками; таким образом, цветочная пыльца осыпается на рыльца в более продолжительный промежуток времени. Чтобы пыльца непременно попала куда следует, можно слегка смочить рыльца жидким раствором сахара в воде. Это имеет ту пользу, что пыльца остается на еще не вполне развитых рыльцах до их совершенного развития и тогда прорастает.
Пыльца, равно как споры и семена, имеет способность сохраняться более или менее продолжительное время; крайний срок в отдельных случаях неизвестен и, вероятно, различен у различных растений и при различных способах хранения. Сберегаемая в капсульке из почтовой бумаги цветочная пыльца бредины (Safe саргеа) сохранила растительную и оплодотворительную способность две недели, – долее, чем могут лежать без утраты всхожести семена этого растения. Пыльца многих других растений может пролежать и долее, не теряя своей способности в прорастанию. Важнейшее пищевое растение Аравии – финиковая пальма, двудомное растение, как наша ива, опыляется (едва ли сознательно) владельцами, которые, по рассказам, пользуются иногда пылью годового возраста. Опыление производится подвешиванием кистей мужских цветов над женскими цветами, так что пыльца сама собою осыпается на последние. На сотню женских растений считается достаточно одного мужского.
Во многих случаях необходимо собирать пыльцу за некоторое время перед оплодотворением, иначе насекомые растащат ее, и, когда наступит время опыления, ничего не останется. Лучшее время для сбора пыльцы – утро, как только растрескиваются пыльники; положенные на стеклянные пластинки или на листок почтовой бумаги, цветки при созревании пыльников в течение дня испускают очень много пыльцы. На простой писчей бумаге или подобном шероховатом материале нельзя собирать цветочную пыльцу; вследствие липкости и мягкости зерен, они пристают к подобным предметам и невозможно удалить их оттуда без повреждения. Для переноски на рыльце такой вытрясенной пыльцы служит мягкая рисовальная кисточка. Лучшим временем для производства опыления считаются утренние часы, от 8 до 10, хотя можно также опылять и вечером; самое жаркое время дня менее удобно.
c) Далее, при опылении, с целью гибридизации, больших древесных растений, например, плодовых деревьев и кустарников, которые пересаживать в горшки невозможно, изолирование цветов от естественного или постороннего опыления составляет весьма существенное условие. Для этой цели весьма удобно применять кисею, сшитую в форме рукава или мешка, который надевают на ветки еще до распускания цветов. Такие вуали с большим успехом употреблял Вихура при его опытах над гибридизацией ив, и я сам, после многочисленных подобных опытов, имел случай убедиться в действительности этой меры. Вуали нисколько не препятствуют принятию искусственного опыления, где таковое вообще возможно, и вполне изолируют цветы от естественного опыления.
Для опытов выбирают отдельные, более сильные ветви и оставляют на них только по нескольку лучших цветов. Удаление тычинок совершается еще до полного распускания цветов, чтобы не последовало естественного опыления. При вскрытии цветков часто случается повреждение лепестков, но это не имеет для оплодотворения вредных последствий, лишь бы рыльца остались нетронутыми. Нельзя вполне надеяться на полное удаление всех тычинок при первой операции; необходимо производить поверку на следующий день (рано и перед опылением), не осталась ли еще в глубине цветка какая-нибудь скрытая тычинка, которая может расстроить весь опыт. Самый прием опыления, равно как и сбор цветочной пыльцы, во всех случаях производятся одинаково.
d) Наконец, самый простой случай тот, когда имеют дело с однодомным растением, например дынею, или двудомным, как, например, хмелем. Тогда требуется только заблаговременное изолирование вуалью и опыление – когда рыльце разовьется до известной степени восприимчивости. Нельзя, однако, вполне рассчитывать на раздельнополость двудомных растений; случается, хотя и весьма редко, что на женских особях встречаются по одиночке тычинки и наоборот, поэтому необходимо тщательно осматривать выбранные для опыта цветки.
e) Признаки удачного опыления. Если опыление удачно, то через день или два результаты уже заметны. Признаками удачного результата служат: почернение и засыхание рыльца, опадение лепестков и увеличение завязи; последний признак, однако, обнаруживается не раньше, как через несколько дней.
Всякая ветка или цветок обозначаются ярлычком, на котором отмечается, чем и когда произведено опыление. Вуаль можно по усмотрению снять или оставить до созревания плодов или семян. Если растение имеет летучие семена, то удобнее оставлять их под покровом вуали, при помощи которой собираются также семена растрескивающихся плодов или стручков.
f) Сеянцы травянистых однолетних, двулетних и многолетних растений скоро достигают возраста плодоношения, и тогда легко убедиться в достоинстве полученного гибрида или помеси. Но у растений древесных пород, особенно у плодовых деревьев, которые требуют очень долгого времени для достижения возраста плодоношения – бывает иначе. Чтобы скорее узнать, какого достоинства полученная новость, можно привить ветвь ее к кроне крупного дичка или маловажного плодового дерева; от такой прививки обыкновенно получаются плоды на 3-м и 4-м году.
Какая особенность производителей будет преобладать в потомстве, трудно определить заранее; прежде думали, что свойства цветка и плодов преимущественно передаются мужским растением; но такое мнение ничем не доказано и едва ли основано на опытах. Вообще можно полагать, что гибриды между естественными видами представляют во всех отношениях среднее между производителями. Всякому на каждом шагу доступен для наблюдения в этом отношении пример: две из самых обыкновенных наших ив – высокая ива, Salix excelsior, происшедшая от ветлы и ломкой ивы (S. alba + S. fragilis), и верболоз, S. acuminata, – от бредины и корзиночной ивы (S. саргеа + S. viminalis или S. с. + S. stipularis), обе прежде принимались за самостоятельные виды; но их легко создать искусственным опылением производителей, чем достаточно доказывается их гибридное происхождение. При всяком опыте гибридизации между данными видами, равно как и между видами вообще (считая их гомозиготными), постоянно получаются одни и те же гибридные формы, лишь иногда с самыми тонкими, едва заметными отклонениями (модификациями). При скрещивании же гибридных или видоизмененных форм (если они гетерозиготны) – дело совершенно другое: от таких растений в потомстве часто получаются уже весьма разнообразные оттенки в различном направлении.
g) Полагают, что гибридные растения, размножаясь половым путем, вскоре возвращаются обратно к одному из производителей; но это мнение справедливо лишь в том случае, если имеет место постоянное оплодотворение со стороны одного из родительских растений; оплодотворенные собственною пылью Аридные растения размножаются половым способом, по крайней мере во многих случаях, так же правильно, как виды. Примером этого служат высокая ива и верболоз, которые совершенно константны при разведении их посевом.
h) Пример многосложных гибридов представляют роды ив, тополей, роз, спирей, вероятно, также и многие другие роды, но они не так тщательно исследованы.
В диком состоянии встречаются растения, являющиеся результатом соединения в одном неделимом четырех видов, но и этот случай еще далек от границы возможных комбинаций.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Размножение плодовых растений
Размножение плодовых растений В практике садоводства большинство растений размножают вегетативно. В основе вегетативного размножения растений лежит их способность восстанавливать весь организм из определенной жизнедеятельной его части – побега, корня, листа,
Размножение
Размножение На практике применяют несколько способов вегетативного размножения вишни: порослью, прививкой, корневыми черенками и зелеными
Размножение
Размножение Размножение ежевики проводят семенами или вегетативно. Семенное размножение ежевики не позволяет сохранить сортовые признаки; их высевают осенью или весной (после хранения в мокром
Размножение вечнозеленых растений
Размножение вечнозеленых растений Для черенков вечнозеленых растений используют побеги с хорошо вызревшей древесиной. В отличие от одревесневших черенков у них сохраняются листья и покой неглубок. А поскольку на черенках есть листья, для предупреждения избыточной
Размножение
Размножение Размножение семенами Из семян выращивают тропические и субтропические кадочные растения. Из крупномеров умеренных широт семенами размножают мирт, цитрусовые.Многие семена, имеющие прочную кожицу, перед высадкой подвергают стратификации. Чаще всего кожицу
Размножение
Размножение Выращивание дыни рассадой В Средней полосе дыню предпочтительнее выращивать через рассаду.За 30 – 35 дней до высадки рассады в грунт, семена дыни высевают в горшочки. Для средней полосы сроки посева приходятся на 20 – 25 апреля. Это самый оптимальный вариант. В
Размножение
Размножение Размножение айвы Айву размножают как семенами, так и вегетативными способами.Семенное размножение айвы. При размножении семенами айва нередко утрачивает свои сортовые признаки. Однако этот способ позволяет обеспечить эффективную акклиматизацию айвы и
Размножение комнатных растений
Размножение комнатных растений Решающим условием успешного выращивания в комнатных условиях плодовых и овощных культур, отличающихся урожайностью, ранним вступлением в плодоношение, устойчивостью к вредителям и болезням, является получение здорового посадочного
Болезни комнатных растений. Симптомы болезней растений
Болезни комнатных растений. Симптомы болезней растений Комнатные растения, как люди и животные, иногда болеют. Болезни комнатных растений имеют свои особенности. Распознать их можно по определенным признакам, симптомам. К сожалению, лечение болезней комнатных растений
Шредер Рихард Иванович
Просмотр ограничен
Смотрите доступные для ознакомления главы 👉